호중구
호중구[1][2](好中球, 영어: neutrophil, neutrocyte) 또는 중성구 또는 이염색 백혈구[3](영어: heterophil) 또는 다형핵 백혈구[4](영어: polymorphonuclear leukocyte)는 포유류에서 가장 많은 비율(40 - 75%)을 차지하는 백혈구이다. 선천 면역에 주요한 역할을 한다. 골수에 있는 줄기세포에서 형성되고, 수명은 짧으며 이동성이 높다. 호중구는 분절형 호중구(segmented neutrophil)와 띠형 호중구(banded neutrophil)로 세분할 수 있다. 호중구는 호산구, 호염기구와 함께 다형핵세포족에 속한다.[5][6][7]

호중구라는 이름은 헤마톡실린과 에오신(H&E 염색)으로 염색하였을 때 조직 혹은 세포의 염색 특성에서 유래하였다. 호염기구가 어두운 파란색으로, 호산구가 밝은 빨강으로 염색되는 반면에 호중구는 그 중간의 분홍색으로 염색된다. 일반적으로 호중구의 핵은 2 - 5개의 엽으로 나뉜다.
호중구는 포식세포의 일종으로 정상적으로는 혈류를 따라 순환한다. 염증(종종 급성 염증에 해당한다)이 시작되는 시기, 특히 세균 감염, 환경에의 노출,[8] 특정 암의 경우에[9][10] 호중구는 가장 빨리 반응하는 세포 중 하나이다. 호중구는 혈관을 통해 이동하여 인터류킨 8, C5a, fMLP, 류코트라이엔 B4 등 화학 신호를 따라 사이질조직을 지나 염증 부위로 이동한다(화학주성). 고름은 호중구가 많아 약간 희고 누르스름한 형태이다.
호중구는 외상 후 수 분 이내에 손상된 부위로 이동하며 급성 염증 반응의 특징적인 세포이다.[11]
특징
편집
호중성 과립구는 말초 혈액 도말 실험에서 평균 지름 12 - 15 μm로 나타난다. 자동 세포계수기에서 순수한 호중구 부유액을 분석하면 평균 지름은 8 - 9μm이다.
림프구와 단핵구 및 기타 백혈구와 달리, 호산구, 호염기구와 함께 호중구는 엽이 있는 핵을 가진 다형핵세포에 속한다. 핵은 특징적인 엽 형태로, 염색질로 서로 연결된다. 호중구는 성숙하면서 핵 분절이 증가하여 과다분절 상태가 된다. 또한 성숙하면서 핵소체가 사라지는 소수의 유핵세포에 속한다.[12]:168 세포질에 있는 골지체는 작고, 미토콘드리아와 리보솜이 드물게 존재하며 조면소포체는 없다.[12]:170 세포질에는 200여 개의 과립이 들어 있고, 그 중 1/3은 아주르친화성 과립이다.[12]:170
수컷과 암컷 개체의 호중구는 약간 다르다. 암컷 호중구의 핵에는 추가적으로 "호중구 북채"라고 알려진 작은 X 염색체 구조가 있다.[12]:174

호중구는 사람의 혈액에서 가장 많은 백혈구이다. 하루에 약 1011개가 생산되고, 모든 백혈구 중 50 - 70%를 차지한다. 표준 호중구 수의 범위는 2.5 - 7.5 x 109/L이다. 아프리카와 중동에서는 호중구 수가 감소하지만 정상 범위에 속한다. 호중구는 다시 분절호중구와 띠형호중구로 세분할 수 있다.
호중구가 혈류를 돌면서 비활성화된 상태일 때는 구형이다. 활성화되면 확실한 형태가 없는, 혹은 아메바 형태로 변하고 항원을 따라다니면서 위족을 뻗을 수 있다.[13]
호중구는 세균보다 정제된 탄수화물, 예를 들어 포도당, 과당, 설탕, 꿀, 오렌지 주스[14] 등을 포식하는 것을 선호한다.[14][15][16] 호중구의 식세포 능력은 단순당을 섭취하였을 때 영향을 받았고,[14] 공복 상태에서는 세균을 잡아먹는 능력이 증가하였다.[14] 일반적인 녹말을 섭취하는 것은 영향을 주지 않았으며, 호중구의 수에는 변화가 없었다.
수명
편집
활성화되지 않고 혈류를 따라 순환하는 사람 호중구의 평균 수명은 5.4일이다.[17] 호중구가 활성화되면 혈관 내피세포에 근접하여 셀렉틴에 의존하여 포착되고 인테그린 의존성 부착으로 이어진다. 혈관을 떠나 조직으로 이동한 호중구는 그곳에서 1 - 2일 생존한다.[18]
호중구는 훨씬 오래 생존하는 다른 포식세포, 즉 단핵구나 대식세포에 비하여 그 수가 훨씬 많다. 병원체(병을 일으키는 미생물이나 바이러스)가 가장 처음 마주치는 것이 대부분 호중구이다. 호중구는 병원체를 소화한 후 대식세포에게 잡아먹히기도 한다. PECAM-1과 포스파티딜세린이 이에 관여한다.
호중구의 수명이 짧은 것이 진화적인 적응이라는 가설도 있다. 숙주 세포 바깥에 존재하는 시간이 길수록 병원체가 체내의 다른 방어 요소에 의하여 파괴될 확률이 높아지므로, 짧은 수명 덕택에 포식세포에 기생하는 병원체의 전파를 최소화할 수 있다. 호중구가 염증 반응 동안 생산하는 항미생물성 물질은 미생물뿐만 아니라 조직을 손상시킬 수 있기 때문에 수명이 짧으면 조직 손상 가능성도 낮아진다.[18]
화학주성
편집호중구는 화학주성을 통하여 감염이나 염증 부위로 이동한다. 세포 표면에 있는 수용체를 통하여 인터류킨 8, 인터페론 감마, C5a, 류코트라이엔 B4 같은 분자의 농도 기울기를 감지하고 길잡이로 이용한다.
호중구에는 특이적인 수용체가 많이 있다. 보체 수용체, 인터류킨과 인터페론 감마에 대한 사이토카인 수용체, 케모카인 수용체, 혈관내피를 감지하고 부착하기 위한 수용체, 렉틴과[19] 단백질에 대한 수용체 및 옵소닌과 결합하는 Fc 수용체 등이 포함된다.[20]
항미생물 기능
편집호중구는 이동성이 높아 감염 부위에 재빨리 모인다. 호중구는 활성화된 혈관내피, 비만세포, 대식세포가 발현하는 사이토카인에 이끌린다. 호중구 또한 사이토카인을 발현하고 방출하여,[21] 그 신호를 받은 다른 세포에 의하여 염증 반응이 증폭됨.
호중구는 면역계의 다른 세포들을 끌어들이고 활성화하기도 하지만, 호중구는 병원체의 침입에 대항하는 최전방에 있는 세포이다. 호중구는 세 가지 방법으로 미생물을 직접 공격한다.[22]
- 식세포작용(섭취)
- 과립 단백질을 포함하여 수용성 항미생물제 방출
- 호중구 세포외덫(neutrophil extracellular trap, NET) 생성
식세포작용
편집호중구는 포식세포로 미생물이 수 있다. 호중구가 표적 물질을 인식하기 위해서는 옵소닌으로 둘러싸이는 과정이 필요한데, 이를 항체 옵소닌화라 한다.[13] 옵소닌화를 통하여 호중구가 미생물을 세포 내로 들이고 파고솜을 형성하면, 활성산소종과 가수분해효소를 파고솜 내로 분비하여 미생물을 죽일 수 있다.
활성산소종을 생산하면서 산소를 소비하는 것을 "호흡폭발"이라 한다. 호흡폭발은 호흡이나 에너지 생산과는 무관하다. 호흡폭발은 활성산소종인 초과산화물을 대량으로 생산하는 NADPH 산화효소가 활성화되는 것과 관련이 있다. 초과산화물은 자발적으로 안정한 상태로 변화하거나 초과산화물 디스뮤테이스(Cu/ZnSOD와 MnSOD)에 의해 분해되어 과산화수소가 되고 녹색 헴 효소 골수세포형과산화효소에 의하여 다시 하이포아염소산(HClO)으로 전환된다. HClO는 호중구가 포식한 세균을 죽일 수 있는 물질이지만, 직접 세균을 처리하는 대신 단백질분해효소를 활성화하는데 필요하다는 연구가 있다.[23]
탈과립
편집호중구는 세 종류의 과립을 통하여 단백질 꾸러미를 방출한다. 이 과정을 탈과립이라 한다. 과립에 들어있는 물질은 항미생물 특성을 가지고 있고, 감염과 싸우는 것을 돕는다.
| 과립 유형 | 단백질 |
| 아주르친화성 과립 (또는 "일차 과립") | 골수세포형과산화효소, 살균/투과성 증가 단백질, 디펜신, 세린 단백질분해효소인 호중구 엘라스테이스와 카텝신 G |
| 특이과립 (혹은 "이차 과립") | 알칼리성 인산분해효소, 라이소자임, NADPH 산화효소, 콜라겐분해효소, 락토페린, 카텔리시딘 |
| 삼차 과립 | 카텝신과 젤라틴분해효소 |
호중구 세포외덫
편집2004년에 브링크만과 동료들은 호중구가 활성화되면 거미줄 구조의 DNA를 방출하며, 이것이 세균을 죽이는 세 번째 기작이라고 보고하였다.[24] 호중구 세포외덫은 염색질과 세린 단백질분해효소로 이루어진 섬유망으로, 미생물을 세포외에서 잡아서 죽인다. 호중구 세포외덫은 항미생물 요소를 국소적으로 집중시켜서 식세포작용을 통한 섭취 없이 미생물에 결합하고, 미생물을 무장해제시켜 없애는 것으로 추정된다. 호중구 세포외덫은 또한 병원체가 더 퍼지지 않도록 물리적 방어벽을 형성한다. 패혈증에서는 혈관에 세포외덫이 형성된다.[25] 호중구 세포외덫이 염증성 질환에서 특정한 역할을 한다는 보고가 있다. 임신과 관계된 염증성 질환인 자간전증에서 호중구가 활성화되고, 따라서 호중구 세포외덫이 어떤 역할을 할 수 있다.[26] 호중구 세포외덫은 인 비트로[27] 및 인 비보[28][29] 실험에서 혈전을 생성하는 효과가 있다고 밝혀졌다.
질병
편집호중구 수가 낮은 질병을 호중구감소증이라 한다. 실험실에서 호중구 수를 계산했을 때 혈액에 보통 2.0×109개/L 이하가 있을 경우 호중구감소증으로 보고한다. 호중구감소증은 여러 가지 원인에 의해 발생할 수 있는데, 선천성 장애나 유전적 질환일 수도 있고, 재생불량성빈혈과 거대적혈모구빈혈, 발작성야간혈색소뇨증과 같이 빈혈에서도 나타나며, 또는 급성백혈병, 다발성골수종같이 골수에 이상이 생겨 생성되는 호중구의 수가 적어 발병하는 경우도 있다. 의약품의 부작용으로 생기기도 하는데, 화학요법으로 인한 경우가 대부분이다.
장티푸스에 감염되었을 경우 다른 세균 감염과 다르게 호중구 수가 감소하는 증상을 동반하며, 그 외에 리케치아균에 의한 발진티푸스에도 호중구가 감소한다. 또한 바이러스에 감염되었을 때도 발병할 수 있으며 이는 바이러스가 직접 호중구를 파괴하기 때문에 발생하는 것으로 생각하고 있다. 그 외에 기생충 감염에도 발병할 수 있는데 말라리아에 감염되었을 경우 감염 초기에는 호중구 수가 다소 증가하는 듯 하다가 열이 진행할수록 그 수가 감소하며, kala-azar와 같은 기생충 감염이 일어났을 때도 감소증이 발생한다. 이렇게 발생한 호중구감소증에 걸린 이는 감염에 매우 취약하다.
알파 1 안티트립신 결핍의 경우에 호중구에서 주요 효소인 엘라스테이스를 저해하는 알파 1 안티트립신이 부족하다. 엘라스테이스가 적절하게 저해되지 않으면 염증이 일어났을 때 조직이 과도하게 손상된다. 만성 폐쇄성 폐질환에서 흔하다.
가족성 지중해열은 pyrin 또는 marenostrin(en) 유전자의 돌연변이로 나타난다. 이 유전자는 주로 호중구에서 발현되어 급성 병기 반응을 일으키고 발열, 관절통, 복막염으로 이어져 결국에는 아밀로이드증을 일으킨다.[30]
호중구 항원
편집현재까지 5개의 호중구 항원(HNA 1-5)이 알려져 있다.[31] HNA-1 항원 세 개(a-c)는 친화성이 낮은 Fc-γ수용체 IIIb(FCGR3B :CD16b)에 있다. 하나만 알려진 HNA-2a은 CD177에 있다. HNA-3 항원계에는 CLT2 유전자(SLC44A2)의 일곱 번째 엑손에 엤는 항원 3a와 3b가 있다. HNA-4와 HNA-5 항원계에는 각각 항원 a와 b가 알려져 있다. 이 항원들은 β2 인테그린에 있는데, HNA-4는 αM 사슬(CD11b)에, HNA-5는 αL 인테그린 단위(CD11a)에 있다.
기타 자료
편집-
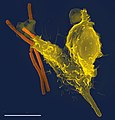
분생자를 쫓아 포식하는 호중구. 30초당 한 프레임으로 2시간 촬영.
-
-
-
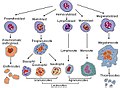 혈액 세포 계보
혈액 세포 계보 -
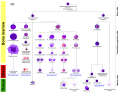 혈액 세포 계보
혈액 세포 계보 -
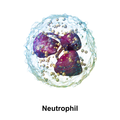 3D 렌더링 한 호중구
3D 렌더링 한 호중구


_diagram_en.svg)

같이 보기
편집각주
편집- ↑ 대한의협 의학용어 사전 https://www.kmle.co.kr/search.php?Search=neutrophil&EbookTerminology=YES&DictAll=YES&DictAbbreviationAll=YES&DictDefAll=YES&DictNownuri=YES&DictWordNet=YES
- ↑ 경북대 치과대학 구강내과 교실 사전 https://www.kmle.co.kr/search.php?Search=neutrocyte&EbookTerminology=YES&DictAll=YES&DictAbbreviationAll=YES&DictDefAll=YES&DictNownuri=YES&DictWordNet=YES
- ↑ 대한의협 의학용어 사전 https://www.kmle.co.kr/search.php?Search=heterophil&EbookTerminology=YES&DictAll=YES&DictAbbreviationAll=YES&DictDefAll=YES&DictNownuri=YES&DictWordNet=YES
- ↑ 대한의협 의학용어 사전 https://www.kmle.co.kr/search.php?Search=polymorphonuclear+leukocyte&EbookTerminology=YES&DictAll=YES&DictAbbreviationAll=YES&DictDefAll=YES&DictNownuri=YES&DictWordNet=YES
- ↑ Witko-Sarsat V, Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L (2000). “Neutrophils: molecules, functions and pathophysiological aspects”. 《Lab Invest》 80 (5): 617–53. doi:10.1038/labinvest.3780067. PMID 10830774.
- ↑ Klebanoff SJ, Clark, RA (1978). “The Neutrophil: Function and Clinical Disorders”. Elsevier/North-Holland Amsterdam. ISBN 0-444-80020-4.
- ↑ Nathan, C (2006년 3월). “Neutrophils and immunity: challenges and opportunities”. 《Nature Reviews Immunology》 6 (March): 173–82. doi:10.1038/nri1785. ISSN 1474-1733. PMID 16498448.
- ↑ Jacobs L, Nawrot TS, de Geus B, Meeusen R, Degraeuwe B, Bernard A, Sughis M, Nemery B, Panis LI (2010년 10월). “Subclinical responses in healthy cyclists briefly exposed to traffic-related air pollution”. 《Environmental Health》 9 (64): 64. doi:10.1186/1476-069X-9-64. PMC 2984475. PMID 20973949.
- ↑ Waugh DJ, Wilson C (2008년 11월). “The interleukin-8 pathway in cancer”. 《Clinical Cancer Research》 14 (21): 6735–41. doi:10.1158/1078-0432.CCR-07-4843. ISSN 1078-0432. PMID 18980965.
- ↑ De Larco JE, Wuertz BR, Furcht LT (2004년 8월). “The Potential Role of Neutrophils in Promoting the Metastatic Phenotype of Tumors Releasing Interleukin-8”. 《Clinical Cancer Research》 10 (15): 4895–900. doi:10.1158/1078-0432.CCR-03-0760. ISSN 1078-0432. PMID 15297389.
- ↑ Cohen, Stephen. Burns, Richard C. (2002). 《Pathways of the Pulp》 8판. St. Louis: Mosby. 465쪽.
- ↑ 가 나 다 라 Zucker-Franklin, Dorothy; Greaves, M.F.; Grossi, C.E.; Marmont, A.M. (1988). 〈Neutrophils〉. 《Atlas of Blood Cells: Function and Pathology》 1 2판. Philadelphia: Lea & Ferbiger. ISBN 0-8121-1094-3.
- ↑ 가 나 Edwards, Steven W. (1994). 《Biochemistry and physiology of the neutrophil》. Cambridge University Press. 6쪽. ISBN 0-521-41698-1.
- ↑ 가 나 다 라 Albert Sanchez, J. L. Reeser, H. S. Lau, P. Y. Yahiku, R. E. Willard, P. J. McMillan, S. Y. Cho, A. R. Magie, and U. D. Register (1973). “Role of sugars in human neutrophilic phagocytosis”. The American Society for Clinical Nutrition. 2013년 9월 8일에 확인함.
These data suggest that the function and not the number of phagocytes was altered by ingestion of sugars. This implicates glucose and other simple carbohydrates in the control of phagocytosis and shows that the effects last for at least 5 hr. On the other hand, a fast of 36 or 60 hr significantly increased (P < 0.001) the phagocytic index.
- ↑ Rubin-Bejerano I, Abeijon C, Magnelli P, Grisafi P, Fink GR (2007년 7월). “Phagocytosis by human neutrophils is stimulated by a unique fungal cell wall component”. 《Cell Host Microbe》 2 (1): 55–67. doi:10.1016/j.chom.2007.06.002. PMC 2083279. PMID 18005717.
- ↑ Kneller, Alyssa (2007). “White blood cells are picky about sugar”. Whitehead Institute. 2013년 8월 9일에 확인함.
- ↑ Pillay J, den Braber I, Vrisekoop N, Kwast LM, de Boer RJ, Borghans JA, Tesselaar K, Koenderman L. (2010년 7월 29일). “In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days”. 《Blood》 116 (4): 625–7. PMID 20410504. 2011년 3월 19일에 원본 문서에서 보존된 문서. 2014년 11월 20일에 확인함.
- ↑ 가 나 Wheater, Paul R.; Stevens, Alan (2002). 《Wheater's basic histopathology: a colour atlas and text》 (PDF). Edinburgh: Churchill Livingstone. ISBN 0-443-07001-6. 2011년 11월 22일에 원본 문서 (PDF)에서 보존된 문서.
- ↑ Geijtenbeek TB, Gringhuis SI (2009년 7월). “Signalling through C-type lectin receptors: shaping immune responses”. 《Nat. Rev. Immunol.》 9 (7): 465–79. doi:10.1038/nri2569. PMID 19521399.
- ↑ Charles N. Serhan, Peter A. Ward, Derek W. Gilroy (2010). 《Fundamentals of Inflammation》. Cambridge University Press. 53–54쪽. ISBN 0-521-88729-1.
- ↑ Ear T, McDonald PP; McDonald (2008). “Cytokine generation, promoter activation, and oxidant-independent NF-kappaB activation in a transfectable human neutrophilic cellular model”. 《BMC Immunol.》 9: 14. doi:10.1186/1471-2172-9-14. PMC 2322942. PMID 18405381.
- ↑ Hickey MJ, Kubes P (2009). “Intravascular immunity: the host–pathogen encounter in blood vessels”. 《Nature Reviews Immunology》 (Nature Publishing Group) 9 (5): 364–75. doi:10.1038/nri2532. PMID 19390567.
- ↑ Segal, AW (2005). “How neutrophils kill microbes”. 《Annu Rev Immunol》 9 (5): 197–223. doi:10.1146/annurev.immunol.23.021704.115653. PMC 2092448. PMID 15771570.
- ↑ Brinkmann Volker, Ulrike Reichard, Christian Goosmann, Beatrix Fauler, Yvonne Uhlemann, David S. Weiss, Yvette Weinrauch, Arturo Zychlinsky (2004년 3월 5일). “Neutrophil Extracellular Traps Kill Bacteria”. 《Science》 (AAAS) 303 (5663): 1532–1535. doi:10.1126/science.1092385. ISSN 0036-8075. PMID 15001782. 2007년 4월 9일에 확인함.
- ↑ Clark SR, Ma AC, Tavener AS, McDonald B, Goodarzi Z, Kelly MM, Patel KD, Chakrabarti S, McAvoy E, Sinclair GD, Keys EM, Allen-Vercoe E, DeVinney R, Doig CJ, Green FHY and Kubes P (2007년 4월). “Platelet Toll-Like Receptor-4 Activates Neutrophil Extracellular Traps to Ensnare Bacteria in Endotoxemic and Septic Blood” (PDF). 《Nature Medicine》 (Nature Publishing Group) 13 (4): 463–9. doi:10.1038/nm1565. ISSN 1078-8956. PMID 17384648.
- ↑ Gupta AK, Hasler P, Holzgreve W, Hahn S (2007년 6월). “Neutrophil NETs: a novel contributor to preeclampsia-associated placental hypoxia?”. 《Semin Immunopathol》 29 (2): 163–7. doi:10.1007/s00281-007-0073-4. ISSN 1863-2297. PMID 17621701.
- ↑ Fuchs TA, Brill A, Duerschmied D, Schatzberg D, Monestier M, Myers DD Jr, Wrobleski SK, Wakefield TW, Hartwig JH, Wagner DD (2010년 9월 7일). “Extracellular DNA traps promote thrombosis”. 《Proceedings of the National Academy of Sciences of the United States of America》 107 (36): 15880–5. doi:10.1073/pnas.1005743107. PMC 2936604. PMID 20798043.
- ↑ Brill A, Fuchs TA, Savchenko A, Thomas, GM, Martinod K, De Meyer SF, Bhandari AA, Wagner DD (2011년 11월 1일). “Neutrophil Extracellular Traps Promote Deep Vein Thrombosis in Mice”. 《Journal of thrombosis and haemostasis : JTH》 10 (1): 136–44. doi:10.1111/j.1538-7836.2011.04544.x. PMC 3319651. PMID 22044575.
- ↑ Borissoff JI, ten Cate, H (2011년 9월). “From neutrophil extracellular traps release to thrombosis: an overshooting host-defense mechanism?”. 《Journal of thrombosis and haemostasis : JTH》 9 (9): 1791–4. doi:10.1111/j.1538-7836.2011.04425.x. PMID 21718435.
- ↑ Ozen, S (2004년 7월). “Familial mediterranean fever: revisiting an ancient disease”. 《European Journal of Pediatrics》 162 (7–8): 449–54. doi:10.1007/s00431-003-1223-x. ISSN 0340-6199. PMID 12751000. 2019년 9월 12일에 원본 문서에서 보존된 문서. 2014년 11월 20일에 확인함.
- ↑ Chu HT, Lin H, Tsao TT, Chang CF, Hsiao WW, Yeh TJ, Chang CM, Liu YW, Wang TY, Yang KC, Chen TJ, Chen JC, Chen KC, Kao CY (2013). “Genotyping of human neutrophil antigens (HNA) from whole genome sequencing data”. 《BMC Med Genomics》 6 (1): 31. PMID 24028078.
외부 링크
편집- (영어) 생체내 호중구(초록색) 영상